Environmental pollution, both visible and invisible, many palliative medications, and increasingly this century the cellular level radiation from wifi devices, are interfering with the capacity of the innate immune system to create the defensive molecules necessary to control intracellular infections
===== Notes and comments =====
Vitamin D, Vitamin D Receptor, and Macroautophagy in Inflammation and Infection
https://tiny.cc/9xtyw
Cool paper, have a look at the references
This paper helps my understanding of innate immune action where I had questions. If it is correct and my understanding of the paper is good, then it looks like general cells use a VDR mediated house cleaning mechanism, autophagy, to deal with intracellular pathogens. This has significant implication to healing if it is correct and if it is turned on by olmesartanMedication taken regularly by patients on the Marshall Protocol for its ability to activate the Vitamin D Receptor. Also known by the trade name Benicar. . I think it means that generally, any infected nucleated cell could clear its intracellular pathogens if the intracellular infection has not blocked VDR creation. It would mean healing could occur without apoptosis of infected cells. That is what it seems to me that the article is implying.
On other points, the paper uses D3 and 1,25D3 interchangeably (because it is the active ligand form of D3), which makes me nervous as researchers usually use D3 and 25D3 interchangeably. I am also not confidant that the papers author knows that murine models are misleading when discussing human VDR.
In general, I am with Chris and would like some help with understanding this paper and its drawbacks.
Makes sense, we've bantered about autophagy in prior threads. The VDR may not be expressed all tissues and red blood cells are not nucleated.
This underscores the need to have s healthy immune syatem.
https://www.reuters.com/article/2011/04/22/us-biogen-tysabri-idUSTRE73L2WN20110422
“Limited data is available beyond four years”
..Trevor..
Cell Microbiol. 2010 Nov;12(11):1648-65. doi: 10.1111/j.1462-5822.2010.01497.x. Epub 2010 Jul 20.
Mycobacterial lipoprotein activates autophagy via TLR2/1/CD14 and a functional vitamin D receptor signalling.
Shin DM, Yuk JM, Lee HM, Lee SH, Son JW, Harding CV, Kim JM, Modlin RL, Jo EK.
Department of Microbiology, Chungnam National University School of Medicine, Daejeon, Korea.
Abstract
In human monocytes, Toll-like receptor (TLR) 2/1 activation leads to vitamin D3Form of vitamin D made in the skin when exposed to light. Also available in fish and meat. This secosteroid is sometimes converted into 25-D. Also known as cholecalciferol and activated 7-dehydrocholesterol.-dependent antimycobacterial activities, but the molecular mechanisms by which TLR2/1 stimulation induces antimicrobial activities against mycobacteria remain unclear. Here we show that TLR2/1/CD14 stimulation by mycobacterial lipoprotein LpqH can robustly activate antibacterial autophagy through vitamin D receptor signalling activation and cathelicidin induction. We found that CCAAT/enhancer-binding protein (C/EBP)-β-dependent induction of 25-hydroxycholecalciferol-1α-hydroxylase (Cyp27b1) hydroxylase was critical for LpqH-induced cathelicidin expression and autophagy. In addition, increases in intracellular calcium following AMP-activated protein kinase (AMPK) activation played a crucial role in LpqH-induced autophagy. Moreover, AMPK-dependent p38 mitogen-activated protein kinase (MAPK) activation was required for LpqH-induced Cyp27b1 expression and autophagy activation. Collectively, these data suggest that TLR2/1/CD14-Ca(2+) -AMPK-p38 MAPK pathways contribute to C/EBP-β-dependent expression of Cyp27b1 and cathelicidin, which played an essential role in LpqH-induced autophagy. Furthermore, these results establish a previously uncharacterized signalling pathway of antimycobacterial host defence through a functional link of TLR2/1/CD14-dependent sensing to the induction of autophagy.
© 2010 Blackwell Publishing Ltd.
PMID: 20560977
Mycobacterial lipoprotein activates autophagy via TLR2/1/CD14 and a functional vitamin D receptor signalling
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2970753/?tool=pubmed
“our findings clearly demonstrate that TLR2/1 signalling regulates antibacterial autophagy pathway through functional vitamin D3 receptor activation and cathelicidin expression in human primary monocytes.”
Nuclear hormone receptors as therapeutic targets.Levi M, Wang X, Choudhury D
Contrib Nephrol 2011; 170() :209-16
Download citation
Affiliation
Division of Renal Diseases and Hypertension, Department of Medicine, University of Colorado Denver, Aurora, Colo., USA.
Abstract
In spite of excellent glucose and blood pressure control, including administration of angiotensin-converting enzyme inhibitors and/or angiotensin II receptor blockers, diabetic nephropathy (DN) still develops and progresses. The development of additional protective therapeutic interventions is, therefore, a major priority. Nuclear hormone receptors regulate carbohydrate metabolism, lipid metabolism, the immune response, inflammation and development of fibrosis. The increasing prevalence of DN has led to intense investigation of the role that nuclear hormone receptors may have in slowing or preventing the progression of renal disease. Several nuclear receptor-activating ligands (agonists) have been shown to have a renal protective effect in the context of DN. This review will discuss the evidence regarding the beneficial effects of the activation of the vitamin D receptor (VDR) and the farnesoid X receptor (FXR) in preventing the progression of DN, and will describe how the discovery and development of compounds that modulate the activity of VDR and FXR may provide potential additional therapeutic approaches in the management of DN.
Also include in “Mechanisms by which…”
Injury enhances TLR2 function and antimicrobial peptide expression through a vitamin D–dependent mechanism
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1784003/?tool=pubmed
An essential element of the innate immune response to injury is the capacity to recognize microbial invasion and stimulate production of antimicrobial peptides. We investigated how this process is controlled in the epidermis. Keratinocytes surrounding a wound increased expression of the genes coding for the microbial pattern recognition receptors CD14 and TLR2, complementing an increase in cathelicidin antimicrobial peptide expression. These genes were induced by 1,25(OH)2 vitamin D3 (1,25D3; its active form), suggesting a role for vitamin D3 in this process. How 1,25D3 could participate in the injury response was explained by findings that the levels of CYP27B1, which converts 25OH vitamin D3 (25D3) to active 1,25D3, were increased in wounds and induced in keratinocytes in response to TGF-β1. Blocking the vitamin D receptor, inhibiting CYP27B1, or limiting 25D3 availability prevented TGF-β1 from inducing cathelicidin, CD14, or TLR2 in human keratinocytes, while CYP27B1-deficient mice failed to increase CD14 expression following wounding. The functional consequence of these observations was confirmed by demonstrating that 1,25D3 enabled keratinocytes to recognize microbial components through TLR2 and respond by cathelicidin production. Thus, we demonstrate what we believe to be a previously unexpected role for vitamin D3 in innate immunity, enabling keratinocytes to recognize and respond to microbes and to protect wounds against infection.
“To further investigate the significance of increased CYP27B1 activation in skin injury, wounds from mice lacking the CYP27B1 enzyme were evaluated for induction of an innate immune response. CD14 is regulated by 1,25D3 (29), and in mouse wounds, CD14 was induced in wild-type animals but not in those lacking CYP27B1 (Figure (Figure5A).5A). In contrast, murine Camp, which is not regulated by 1,25D3 (7), was increased in wounds in both wild-type and CYB27B1-deficient animals”
In VivoA type of scientific study that analyzes an organism in its natural living environment. Activation of the Intracrine Vitamin D Pathway in Innate Immune Cells and Mammary Tissue during a Bacterial Infection.
https://www.ncbi.nlm.nih.gov/pubmed/21124742
Numerous in vitro studies have shown that toll-like receptor signaling induces 25-hydroxyvitamin D(3) 1α-hydroxylase (1α-OHase; CYP27B1) expression in macrophages from various species. 1α-OHase is the primary enzyme that converts 25-hydroxyvitamin D(3) to 1,25-dihydroxyvitamin D(3) (1,25(OH)(2)D(3)). Subsequently, synthesis of 1,25(OH)(2)D(3) by 1α-OHase in macrophages has been shown to modulate innate immune responses of macrophages. Despite the numerous in vitro studies that have shown 1α-OHase expression is induced in macrophages, however, evidence that 1α-OHase expression is induced by pathogens in vivo is limited. The objective of this study was to evaluate 1α-OHase gene expression in macrophages and mammary tissue during an in vivo bacterial infection with Streptococcus uberis. In tissue and secreted cells from the infected mammary glands, 1α-OHase gene expression was significantly increased compared to expression in tissue and cells from the healthy mammary tissue. Separation of the cells by FACS9 revealed that 1α-OHase was predominantly expressed in the CD14(+) cells isolated from the infected mammary tissue. The 24-hydroxylase gene, a gene that is highly upregulated by 1,25(OH)(2)D(3), was significantly more expressed in tissue and cells from the infected mammary tissue than from the healthy uninfected mammary tissue thus indicating significant local 1,25(OH)(2)D(3) production at the infection site. In conclusion, this study provides the first in vivo evidence that 1α-OHase expression is upregulated in macrophages in response to bacterial infection and that 1α-OHase at the site of infection provides 1,25(OH)(2)D(3) for local regulation of vitamin D responsive genes.
I have attached a recent paper that discusses the previously unrecognized function of prions as anti-microbial agents, sharing certain similarities with archetypal anti-microbial peptides like LL-37. I seem to recall you mentioning that you had attended a presentation in Karolinska that discussed how “prion disease” could only exist against the backdrop of a TH1 state. Certainly, the observation that many prions are “infectious” can now be explained by your model. It would appear that prion production may simply be the body's reaction to infection, rather than the cause of “prion disease.”
From the paper:
“As mentioned above, prion protein is not only confined to the nervous system, but instead ubiquitously found in many other cells and tissues, and the physiological role for this protein are still enigmatic. In a previous study, it was reported that human keratinocytes express PrPc in vitro and during inflammatory skin disease [18]. Although that previous work was focusing on prion infectivity routes, our current findings on increased expression of PrP during wounding, together with the observation of its antimicrobial activity, clearly indicate that PrPs could have a previously undisclosed role in host defense. In this context, experiments with PrP deficient animals in infection models should be valuable in order to further delineate a possible role of PrP in innate defense.”52)
“Notably, PrP bound LPS similarly to human LL-37.”
Dan Rudderow
Some AMPs have activity against certain species of antibiotic-resistant bacteria.(11557478)
Hydrogen sulfide: a marker of inflammation:
https://www.eurekalert.org/pub_releases/2010-08/tpco-reg082010.php
https://onlinelibrary.wiley.com/doi/10.1111/j.1749-6632.2010.05556.x/full
In a 2008 paper, Liu lists the different pattern recognition receptors in humans.53)
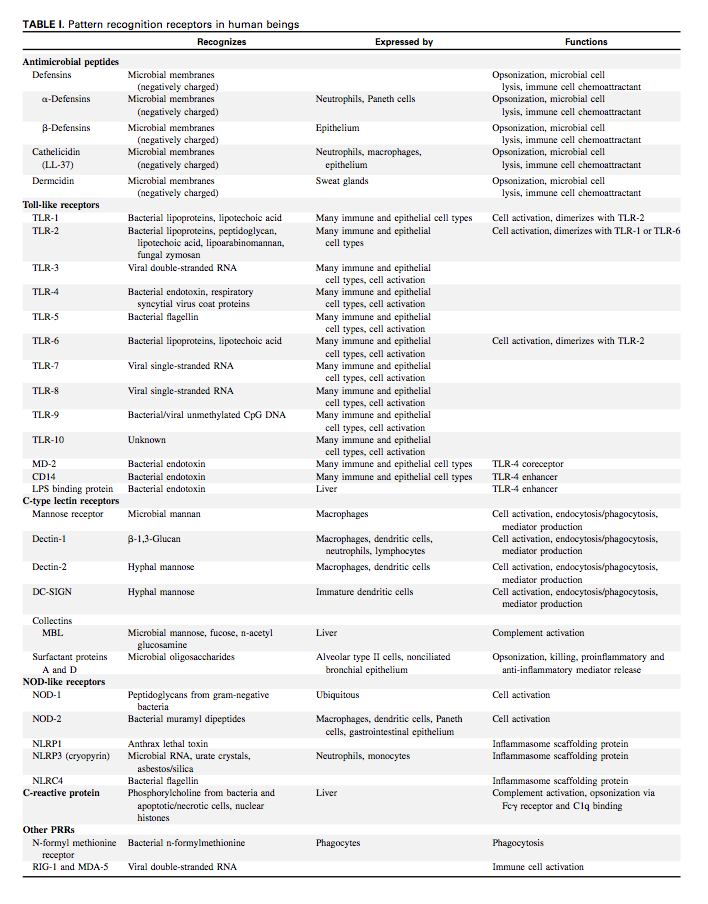
Clin Biochem. 2010 Jul 23. [Epub ahead of print]
Correlation of a multi-cytokine panel with clinical disease activity in patients with rheumatoid arthritis.
Milman N, Karsh J, Booth RA.
Faculty of Medicine, University of Ottawa, 451 Smyth Rd., Ottawa ON, Canada K1H 8M5; Department of Medicine, Division of Rheumatology, 1967 Riverside Dr. Ottawa ON, Canada K1H 7W9.
Abstract
OBJECTIVE: Explore the potential use of a cytokine panel as biochemical markers of disease activity in rheumatoid arthritis (RA) patients. DESIGN AND METHODS: 57 adult RA patients were assessed using five validated clinical disease activity tools: Health Assessment Questionnaire (HAQ), standard 28-joint Disease Activity Score (DAS28), DAS28 using C-reactive protein (DAS28-CRP), Clinical Disease Activity Index (CDAI), and Simple Disease Activity Index (SDAI). Plasma cytokine levels (IL-2, IL-4, IL-6, IL-8, IL-10, VEGF, IFN-gamma, TNF-alpha, IL1alpha, IL1beta, MCP1, and EGF) were measured in 47 of the 57 patients and correlated with clinical indicators. RESULTS: We found significant correlations between plasma levels of IL-6 and all clinical measures of disease activity; Spearman coefficients (p values) were: HAQ: 0.347(0.017); DAS28: 0.409(0.005); DAS-CRP: 0.378(0.011); CDAI: 0.312(0.033); SDAI: 0.310(0.039); ESR: 0.448(0.002); and CRP: 0.513(0.001). IFN-gamma also correlated with DAS-CRP: 0.309(0.039) and SDAI: 0.301(0.044). Furthermore, the levels of IL-6 and IFN-gamma increased significantly with worsening disease, as defined by the European League Against Rheumatism (EULAR) classification of disease activity. CONCLUSION: A significant correlation between plasma levels of IL-6 and clinical disease activity in patients with RA suggests a future role of IL6 as a disease activity marker. Copyright © 2010. Published by Elsevier Inc.
PMID: 20655893
Plasma cytokine fluctuations over time in healthy controls and patients with fibromyalgia.
Togo F, Natelson BH, Adler GK, Ottenweller JE, Goldenberg DL, Struzik ZR, Yamamoto Y.
Pain & Fatigue Study Center, Department of Neurosciences, UMDNJ-New Jersey Medical School, 1618 ADMC, 30 Bergen Street, Newark, NJ 07103, USA.
We examined the pattern of cytokine secretion across the 24-hr day for women with widespread pain and tenderness having the diagnosis of fibromyalgia (FM) and matched healthy controls. Subjects were given time to habituate to being in a clinical research laboratory environment and then were sampled for cytokines without their being disturbed for a 24-hr period including an 8-hr sleep period. Cytokine levels were uniformly low but characterized by bursts of secretion. Bursting occurred either in singlets or in doublets with a range from 88 to 131 mins between doublet bursts. There was an element of synchronization of these bursts with most occurring at the beginning of sampling. FM patients showed a shift to increased IL-10 in the nighttime compared to controls. The relation between this anti-inflammatory cytokine to the pro-inflammatory cytokines studied also differed between groups: FM patients showed an increased ratio of IL-10 burst amplitude to that of pro-inflammatory cytokines IL-1beta, IL-8, and TNF-alpha. We interpret this to indicate a skew away from the normal balance favoring pro-inflammatory cytokines in controls toward one favoring an anti-inflammatory response in FM. These changes toward anti-inflammatory predominance in FM may explain their common complaint of disturbed sleep because these cytokines are known to disrupt sleep.
PMID: 19064941 [PubMed - indexed for MEDLINE]
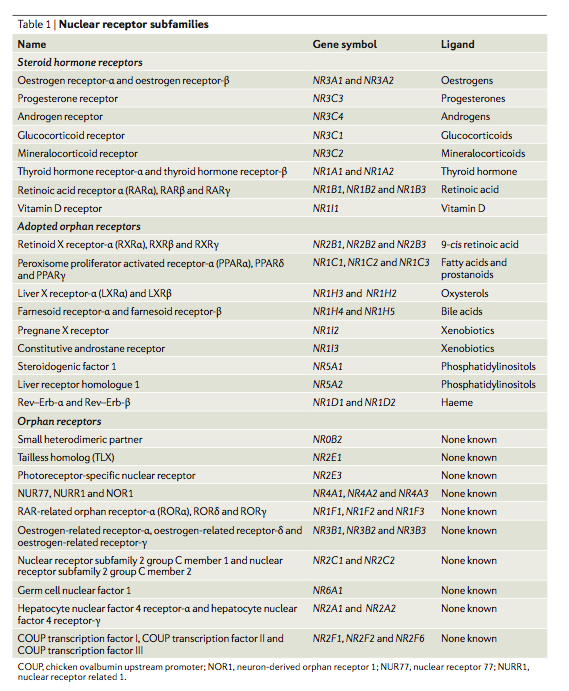
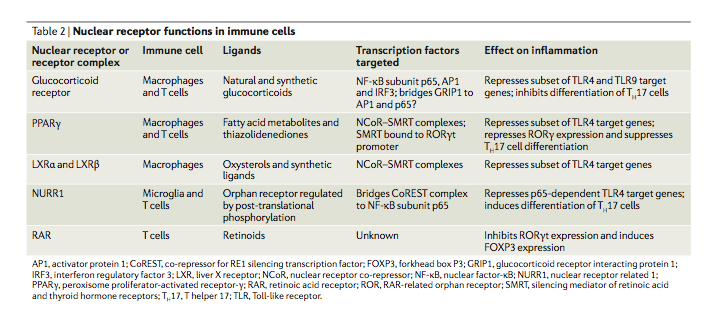
Nuclear receptor transrepression pathways that regulate inflammation in macrophages and T cells54)
===== References =====



